the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 01 Aug 2019
| 01 Aug 2019
Modelling future hydroclimatic effects on the Coregonus migratorius spawning migration in the Selenga River and Lake Baikal
Tatiana Millionshchikova
Sergey Chalov
Regional climate change affects the flow conditions in river basins which can impact the health of aquatic ecosystems. Potential impacts of future climate scenarios on Coregonus migratorius spawning migration in the Selenga River were assessed. A regional process-based hydrological model was used to reproduce the historical trends in the annual flow and assess its future changes under several climate change scenarios. Annual flow projections were used to identify preferential river reaches for spawning activity of the Arctic cisco (Coregonus migratorius), based on the significant negative correlation of spawning activity with the Selenga River streamflow. The applied methodology shows that the projected decline in runoff of 10 % to 25 % in XXI century may result in shifting of the spawning locations further upstream of the Ulan-Ude city, a local “pollution hotspot”.
- Article
(2212 KB) - Full-text XML
- BibTeX
- EndNote





In natural ecosystems, environmental abiotic drivers may play a crucial role in the development and survival of living organisms (Poff et al., 1997; Poff and Zimmerman, 2010; Stagl and Hattermann, 2016). In riverine ecosystems, the reproduction, foraging, maturation and recruitment of aquatic species are dependent on flow and temperature regime, both of which are vulnerable to natural and anthropogenic impacts (Gillenwater et al., 2006; Goto et al., 2015; Stagl and Hattermann, 2016; Wang et al., 2019). The health of aquatic ecosystems is strongly affected by catchment-scale human activities, such as dam construction (Fan et al., 2015; Wang et al., 2019; Wu et al., 2019), irrigation (Bączyk et al., 2018; Nhiwatiwa et al., 2017) and changes in hydro-climatic regime (Goode et al., 2012; Johnson et al., 2009; Statham, 2012). The observed changes in river flow due to these impacts can be evaluated using indicators of alteration in a riverine system (Gao et al., 2009; Richter et al., 1996). However, a commonly applied index for a basic assessment of the alteration of an aquatic system is the average annual streamflow discharge (Bragg et al., 2005).
The most interacting with the river flow are migratory fish species, such as salmonids, spawning in fresh water. The basic water quantity requirements for salmonid fish includes: flows (at appropriate times of the year), water depths, and flow velocities for spawning fish (Hendry et al., 2003). A reduction in flow causes a reduction in water levels and velocities, which is turn decreases available habitat and reduces habitat diversity (Dewson et al., 2007). Low flow conditions can be also attributed to other negative impacts such as increased sediment deposition and decrease in water quality. Numerous studies have used experimental flumes in the laboratory to examine the critical swimming speeds and optimal longitudinal velocities on the microhabitat level, including how velocity and turbulence influence on the variation in swimming behaviour (see Hockley et al., 2014). Whereas almost no studies were attributed to the effects of velocity to fish migration patterns of migratory fishes.
In recent studies a common methodology for assessment of future impacts of climate change on river flow and subsequently on the ecological state of the catchment system is involving a hydrological model forced by expected climate variables to simulate future flows and other ecohydrological indicators (Döll and Zhang, 2010; Gelfan et al., 2017; Moreido and Kalugin, 2017; Papadaki et al., 2016; Schneider et al., 2013; Shrestha et al., 2014; Thompson et al., 2014). Several studies have previously addressed the impact of future climate change on conditions of aquatic environments, such as anadromous fish morphology (Andres et al., 2019) and spawning success (Fenkes et al., 2016).
The present study examines hydroclimatic effects due to climate change on the seasonal spawning migration of Arctic cisco (Coregonus migratorius) in the Selenga Selenga River, which is the largest tributary of Lake Baikal and provides nearly a half of the annual inflow of water to the lake. The observed climate change impacts on the annual flow and regional hydrology (Karthe et al., 2014, 2017; Kasimov et al., 2017; Frolova et al, 2017) have a high potential to impair aquatic ecosystem functions and spawning migration of Coregonus migratorius. The Selenga River remains one of the rivers in the world free of large flow alteration projects (e.g. dams and water abstractions) of the comparable area (Frolova et al., 2017; Törnqvist et al., 2014).
Our assessment involved the construction of a regional-scale hydrological model to reproduce the current flow conditions of the Selenga River, forcing it with the future scenarios of climate change and subsequently applying the obtained results to a regression-based model the spawning migration of Coregonus migratorius.
The Selenga River is a large transboundary river originating in Mongolia and flows across the Republic of Buryatia (Russia) to lake Baikal where it forms a large delta (Fig. 1). The Selenga River is 1024 km in length and drains an area of 450 000 km2. Total annual flow into the lake is 29 km3 and the highest flows occur in August–September. The Selenga River provides spawning habitat for the Coregonus migratorius (see Sect. 3.1). For the last two decades, annual flow of the Selenga River has decreased significantly (Frolova et al., 2017). With its diverse catchment flow generation conditions, ranging from dry steppe to mountainous taiga, and occasional permafrost patterns, the fate of the Selenga River and its ecosystem health under the expected climate change (Karthe et al., 2017; Törnqvist et al., 2014) is of crucial interest.
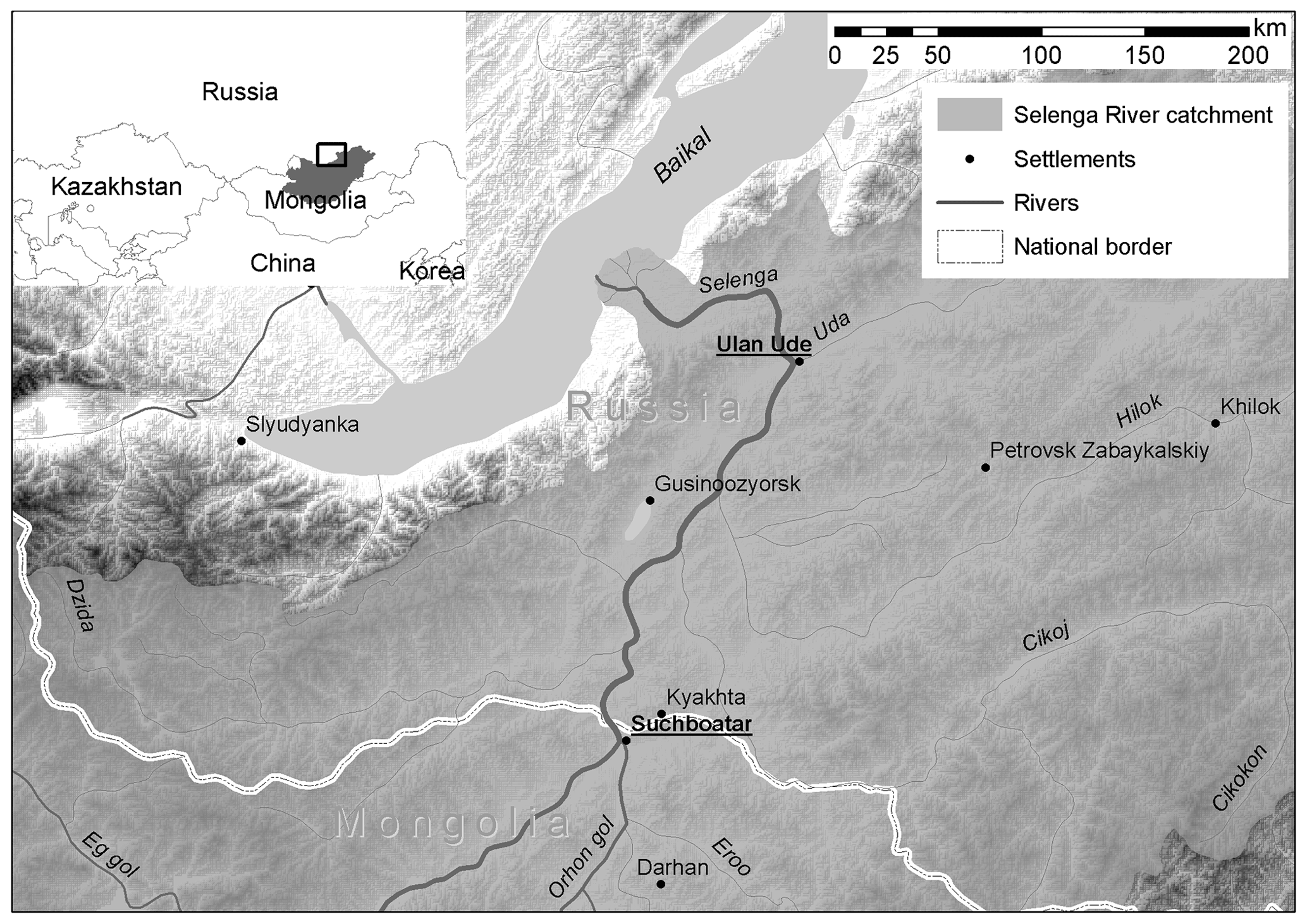
Figure 1The location of the study region.
3.1 Coregonus migratorius migration behavior assessment in current conditions
The Arctic cisco (Coregonus migratorius) is a migratory species of the Salmonidae family foraging in Lake Baikal and spawning in the tributaries. The Selenga and Upper Angara rivers contain the highest populations of Arctic cisco. estimated population of 1.0–3.7 million followed by spawning populations in the Barguzin and Bolshaya rivers (population around 0.1–0.7 million) (Bazov and Bazova, 2015).
Annual spawning migration occurs between the end of August and the end of October. According to multi-year monitoring programs there is a strong negative relationship () between river stage and spawning migration distance during the spawning period (Bazov and Bazova, 2015; Sokolov and Kalyagin, 2001; Voronov, 1993). Preferable spawning conditions for Coregonus species include gravel bed (Roseman et al., 2007), low channel slope and flow velocities between 0.8–2.8 m s−1 (Bazov and Bazova, 2016). There has been no evidence in the local habitat studies of Coregonus migratorius spawning on inundated floodplains during high flows. During the spawning period, water temperature can vary between +9 and +20 ∘C and there is no significant correlation between temperature and spawning activity (Bazov and Bazova, 2015). This is likely due to the tendency of Coregonus migratorius to avoid additional energy loss due to overcoming the increased stream velocity.
Coregonus migratorius typically migrate 20 to 450 km upstream from the delta to spawn (Bazov and Bazova, 2015). A major industrial centre Ulan-Ude (capital of Buryatia Republic) is located 153 km upstream from the delta. Industrial activity represents a pollution hot spot at this location (Karthe et al., 2017). Accordingly, spawning activity is most pronounced in a reach river 140 km from the delta with an average distance of 110 km (Bazov and Bazova, 2010). During periods of low flow spawning activity can occur over a range of 175 to 450 km from the delta with an average distance of 240 km. The analysis of the dependency between the spawning distance and the mean water stage at the Kabansk gage in September showed a clear negative correlation of −0.77 (Bazov and Bazova, 2015), implying that the higher the stage, spawning activity occurs closer to the delta.
Bazov and Bazova (2015) outline the two types of Coregonus migratorius spawning behaviour:
-
“High-stage type” with the majority of spawning occurring downstream of Ulan-Ude, with the average distance of 110±2 km, characterized by high water stage at Kabansk gage cm;
-
“Low-stage type” with the average spawning distance of 240±26 km and water stage of 244±15 cm.
3.2 The Selenga river flow modelling
ECOMAG modelling software was used to model current and assess future flow conditions of the Selenga river. The ECOMAG (ECOlogical Model for Applied Geophysics) is a spatially-distributed process-based hydrological model that considers hydrological processes such as rainfall/snowfall canopy interception, snow accumulation and melt, soil freezing and thawing, water infiltration into unfrozen and frozen soil, evapotranspiration, thermal and water regime of soil, overland, subsurface, ground and river flow with a daily time step (Motovilov et al., 1999; Motovilov, 2016). The model is capable of reproducing streamflow discharge at any point in the river network and is driven by time series data based on daily air temperature, humidity, and precipitation intensity. The ECOMAG model has been applied to the Selenga river basin to assess the future impact of the regional climate change on annual flow using EWEMBI reanalysis dataset (Lange, 2016) as input (Gelfan and Millionschikova, 2018).
The model parameters were calibrated against the observed daily streamflow discharge data at the most downstream hydrometric gauge located at Kabansk (Fig. 1) to reproduce the observed trends in annual flow of the Selenga River. The model showed good agreement between the measured and the calculated annual flow with the following efficiency estimates. For the calibration period (2000–2013) NSE = 0.82, r=0.93 and BIAS = 0.22 %, for the validation period (1986–1999) NSE = 0.77, r=0.88 and BIAS = 4.4 %.
3.3 Assessment of the Selenga flow and Coregonus migratorius spawning conditions under future climate changes
To assess annual flow dynamics in the Selenga river under future climate changes output from the General Circulation Models was used (GCMs) from the ISIMIP2: HadGEM2-ES, IPSL-CM5A-LR, MIROC-ESMCHEM, GFDL-ESM2M, which operated under different conditions of the expected external atmospheric radiation impacts (RCP-scenarios – Representative Concentration Pathways; IPCC, 2014). The original GCMs data were interpolated to a grid and bias-corrected on the continental scale (the bias-correction procedure concept is described in Hempel et al., 2013). The expected changes in the external parameters of the climate models – the scenarios of anthropogenic greenhouse gases' emissions – were introduced according to the radiation impact level for each of the four RCPs expected by 2100: RCP 2.6, RCP 4.5, RCP 6.0, RCP 8.5 (2.6, 4.5, 6.0, 8.5 W m−2, respectively; IPCC, 2014). Based on the four GCMs forced by four RCPs, an ensemble of possible climate projections for the Selenga River basin in the XXI century was obtained with correction factors that have been determined for the historical low-flow period (1996–2005). The correction commonly consisted of minimizing the bias-error of GCM-based calculations on the regional scale (Gelfan et al., 2015) compared with the EWEMBI-based calculations.
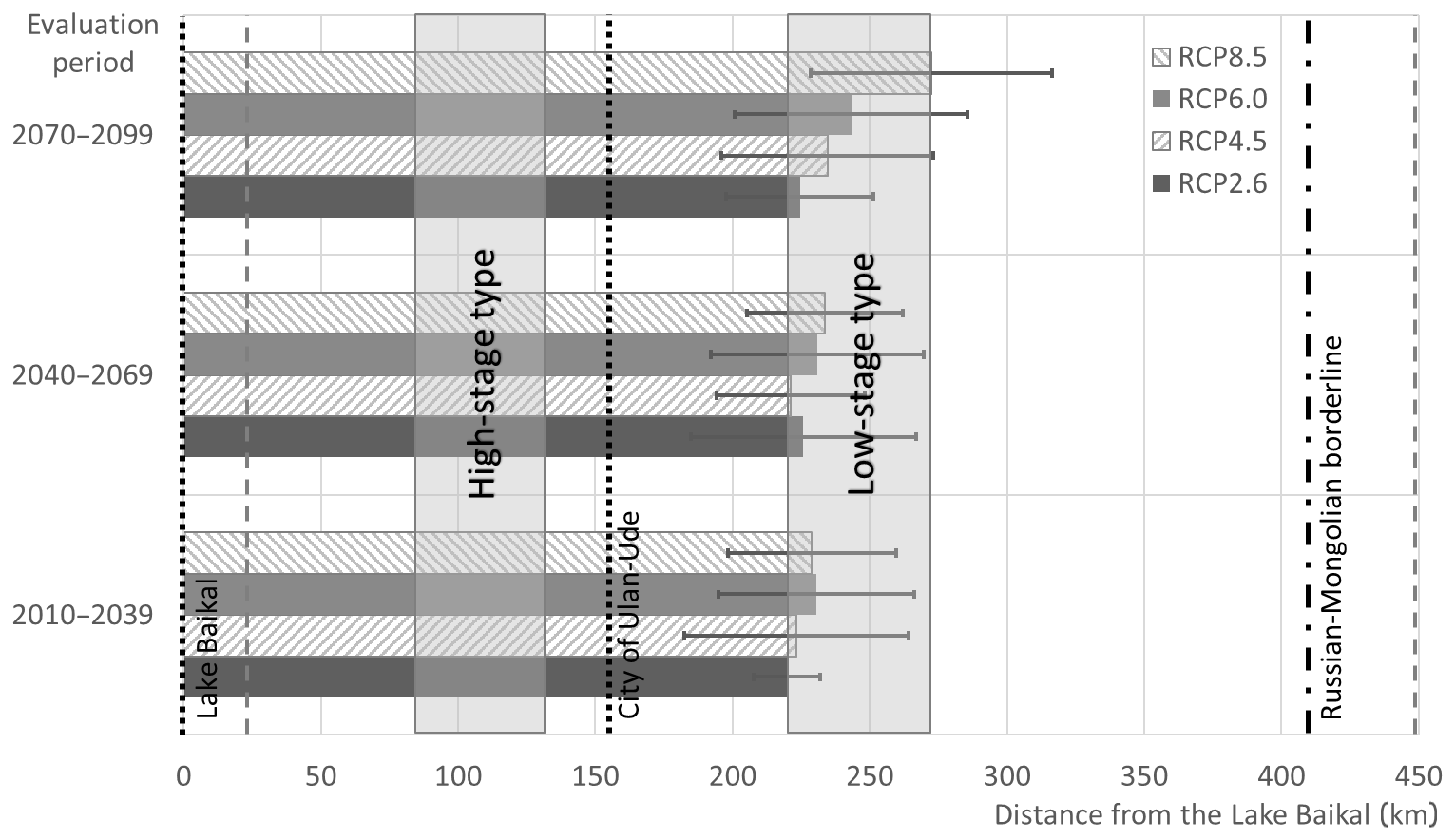
Figure 2Projections of the Baikal omul spawning distance range in the XXI century. Horizontal bars show mean projected distance for different periods, whiskers show inter-model variance. Vertical grey dashed lines denote the historical minimum and maximum spawning ground location, the shaded areas show the two types of the migration distance from the assessment of 1984–2013 (see Sect. 3.1). Vertical black dotted line marks the location of the Ulan-Ude, black dash-dotted line shows the interstate borderline location.
This approach is unsuitable for intra-annual flow projections (Moreido and Kalugin, 2017) due to large uncertainties in the annual cycle described by the GCMs. Hence we used a linear regression model to change between the modelled annual flows and the September streamflow. The correlation coefficient between these variables is 0.91, which allows for such a transition. Next, the average streamflow value in September was converted into the mean stage value with the stage-discharge curve, established using long term data from the Kabansk gauging station. The analysis of the observed mean stage of the Selenga River in September and the calculated using the described procedure showed a good agreement with correlation coefficient r=0.95.
To predict Coregonus migratorius spawning features under future hydrological conditions we used a conceptual modelling approach. Accordingly, we employed a regression model (Bazov and Bazova, 2015) which links hydrological controls of the migration extent using relationship between distance of Baikal Coregonus migratorius spawning sites from the delta with the average water level of the Selenga river at most downstream gage in Kabansk in September.
4.1 Streamflow modelling for XXI century
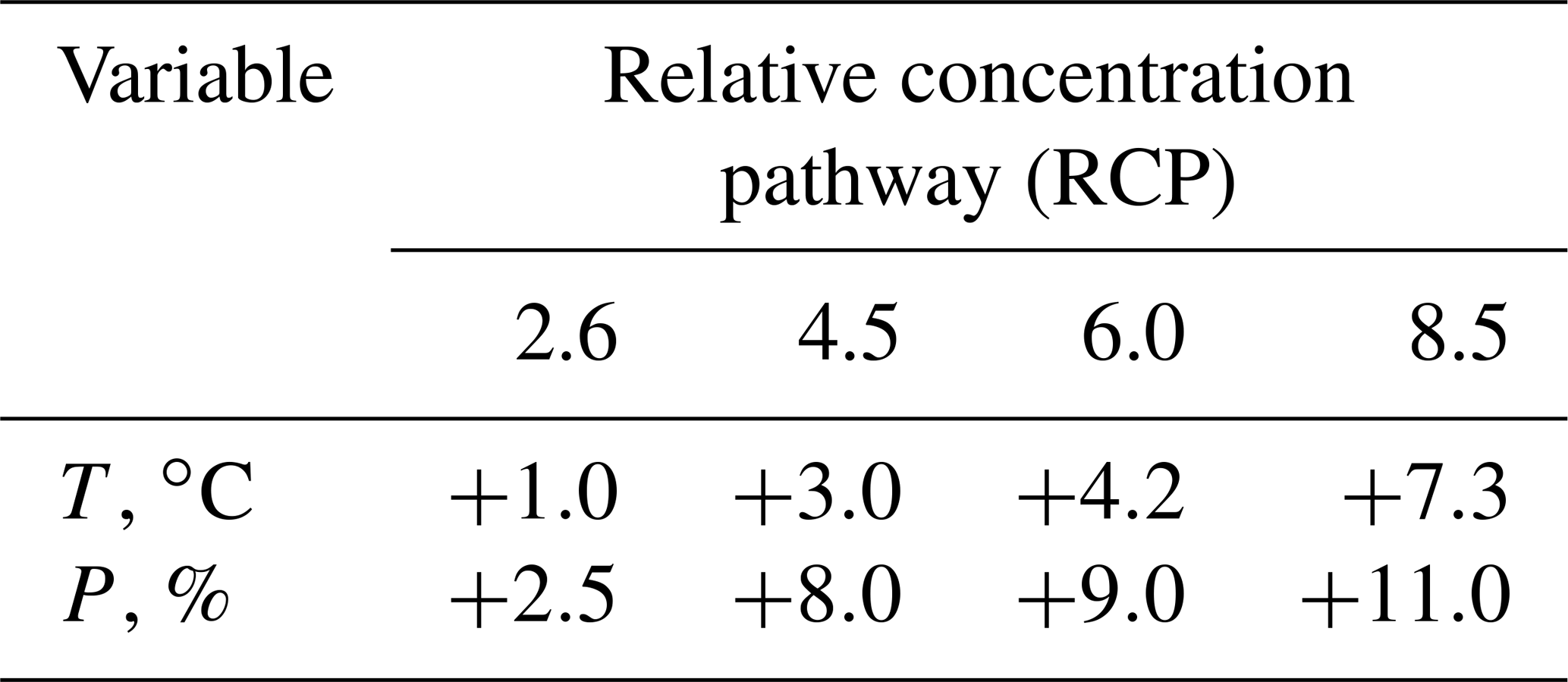
The future regional climate trends for the Selenga river based on the ensemble of four climate models' projections throughout the XXI century were obtained. The ensemble-averaged projections were used to assess the anomalies of air temperature (∘C) and precipitation (%) over three thirty-year-long periods for the XXI century (2010–2039, 2040–2069, 2070–2099) as compared to the historical period of 1996–2005. Based on these results, the trend in the anomaly of the average annual air temperature in the Selenga Basin in the XXI century under different RCP-scenarios increased over time (Table 1). Accordingly, the trend in the anomaly of the average annual precipitation under different scenarios is projected to be positive (Table 1).
Table 1Ensemble-averaged projections of air temperature (∘C) and precipitation (%) anomalies for the XXI century in the Selenga river basin.
Using these projections as inputs to the hydrological model, we simulated flow changes in the Selenga River basin. According to all climate projections, mean annual flow will decrease and this will be more pronounced in the second half of the XXI century. Under the RCP 8.5 scenario flow will decrease by 25 % relative to the historical period. It is assumed that the low-flow conditions into Lake Baikal will remain throughout the XXI century.
4.2 Assessment of the future alteration of the Baikal omul spawning migration
Using the obtained mean annual Selenga River streamflow discharge for future periods at the Kabansk gauge, we calculated the mean stage for these periods and subsequently – the possible alteration of the Baikal omul spawning distance using the methodology described in Sect. 3.1. Figure 2 shows the comparison of the current and the expected conditions of spawning grounds' location expressed as the distance upstream from the Selenga River delta.
The assessment of spawning locations based on the control catch data of 1984–2013 (Bazov and Bazova, 2015) showed that 25 % to 75 % of spawning grounds are located between 110 and 250 km from the delta. Our results show that according to the expected decrease in the Selenga River flow, the spawning migration distance will propagate further upstream. In the first period of 2010–2039 the distance assessed under different RCPs is projected at 219 to 230 km, in the second period – between 221 and 233 km, in the third – between 225 and 272 km. The inter-RCP and inter-model variance increase between the periods, with the most significant difference in the third period of 2070–2099 (Fig. 2).
On average, all the spawning grounds are projected to be located upstream of the Ulan-Ude city, which is ecologically favorable. Most of the projected distances fall into the “low-stage type” of the spawning behavior.
A modelling approach was used to assess the effect of climate change on streamflow and hydrological conditions for Coregonus migratorius spawning. Regional climate change projections from Global Circulation Models operating under different greenhouse gas forcing was determined and a regional hydrological model projecting future climate conditions was used with a regression-based spawning ground location model. The resulting projections show that annual flow of the Selenga River will decrease from 10 % to 25 % according to different climate change scenarios. These changes in river flow indicate that Coregonus migratorius will propagate further upstream of the Selenga River delta to more favorable spawning locations. Further investigations should include a detailed assessment of site scale physical, chemical and biological factors contributing to spawning success at the site, reach and watershed scale.
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
VM and SC designed the study. TM performed the modelling and climate change assessment and wrote the corresponding section. VM performed the final assessment and wrote the rest of the paper. VM and SC made final approval for the publication.
The authors declare that they have no conflict of interest.
This article is part of the special issue “Land use and climate change impacts on erosion and sediment transport”. It is a result of the ICCE Symposium 2018 – Climate Change Impacts on Sediment Dynamics: Measurement, Modelling and Management, Moscow, Russia, 27–31 August 2018.
This research has been supported by the Russian Fund for Basic Research (project 17-29-05027) and State scientific assignment #0147-2019-0003 (reg. #AAAA-A18-118022090105-5).
Andres, K. J., Chien, H., and Knouft, J. H.: Hydrology induces intraspecific variation in freshwater fish morphology under contemporary and future climate scenarios, Sci. Total Environ., 671, 421–430, https://doi.org/10.1016/j.scitotenv.2019.03.292, 2019.
Bączyk, A., Wagner, M., Okruszko, T., and Grygoruk, M.: Influence of technical maintenance measures on ecological status of agricultural lowland rivers – Systematic review and implications for river management, Sci. Total Environ., 627, 189–199, https://doi.org/10.1016/j.scitotenv.2018.01.235, 2018.
Bazov, A. V. and Bazova, N. V.: The monitoring assessment of the Baikal omul egg laying conditions in the Selenga river spawning grounds (lake Baikal basin), in: Biology, biotechnics of the breeding and resources conditions of the Coregonidae fish: Proceedings of the 7th International Applied Scientific Workshop, 16–18 February 2010, Tyumen, Russia, 70–74, Gosrybcentr, 2010 (in Russian).
Bazov, A. V. and Bazova, N. V.: Characteristics of spawning migration of the Baikal | Omul in the Selenga river according to long-term observations (1920–2014), Bull. Fish. Sci., 2, 18–28, 2015.
Bazov, A. V. and Bazova, N. V.: Selenga population of the Baikal omul: the past, the present and the future, Baikal Scientific Centre of the Siberian Branch of the Russian Academy of Sciences, Ulan-Ude, Russia, 2016.
Bragg, O. M., Black, A. R., Duck, R. W., and Rowan, J. S.: Approaching the physical-biological interface in rivers: a review of methods for ecological evaluation of flow regimes, Prog. Phys. Geogr. Earth Environ., 29, 506–531, https://doi.org/10.1191/0309133305pp460ra, 2005.
Dewson, Z. S., James, A. B. W., and Death, R. G.: A review of the consequences of decreased flow for instream habitat and macroinvertebrates, J. North Am. Benthol. Soc., 26, 401–415, https://doi.org/10.1899/06-110.1, 2007.
Döll, P. and Zhang, J.: Impact of climate change on freshwater ecosystems: a global-scale analysis of ecologically relevant river flow alterations, Hydrol. Earth Syst. Sci., 14, 783–799, https://doi.org/10.5194/hess-14-783-2010, 2010.
Fan, H., He, D., and Wang, H.: Environmental consequences of damming the mainstream Lancang-Mekong River: A review, Earth-Sci. Rev., 146, 77–91, https://doi.org/10.1016/J.EARSCIREV.2015.03.007, 2015.
Fenkes, M., Shiels, H. A., Fitzpatrick, J. L., and Nudds, R. L.: The potential impacts of migratory difficulty, including warmer waters and altered flow conditions, on the reproductive success of salmonid fishes, Comp. Biochem. Phys. A, 193, 11–21, https://doi.org/10.1016/j.cbpa.2015.11.012, 2016.
Frolova, N. L., Belyakova, P. A., Grigoriev, V. Y., Sazonov, A. A., Zotov, L. V., and Jarsjö, J.: Runoff fluctuations in the Selenga River Basin, Reg. Environ. Change, 17, 1965–1976, https://doi.org/10.1007/s10113-017-1199-0, 2017.
Gao, Y., Vogel, R. M., Kroll, C. N., Poff, N. L., and Olden, J. D.: Development of representative indicators of hydrologic alteration, J. Hydrol., 374, 136–147, https://doi.org/10.1016/j.jhydrol.2009.06.009, 2009.
Gelfan, A. N. and Millionschikova, T. D.: Validation of a Hydrological Model Intended for Impact Study: Problem Statement and Solution Example for Selenga River Basin, Suppl. 1, Water Resour., 45, S90–S101, 2018.
Gelfan, A. N., Semenov, V. A., Gusev, E., Motovilov, Y., Nasonova, O., Krylenko, I., and Kovalev, E.: Large-basin hydrological response to climate model outputs: uncertainty caused by internal atmospheric variability, Hydrol. Earth Syst. Sci., 19, 2737–2754, https://doi.org/10.5194/hess-19-2737-2015, 2015.
Gelfan, A. N., Gustafsson, D., Motovilov, Y., Arheimer, B., Kalugin, A., Krylenko, I., and Lavrenov, A.: Climate change impact on the water regime of two great Arctic rivers: modeling and uncertainty issues, Clim. Change, 141, 499–515, https://doi.org/10.1007/s10584-016-1710-5, 2017.
Gillenwater, D., Granata, T., and Zika, U.: GIS-based modeling of spawning habitat suitability for walleye in the Sandusky River, Ohio, and implications for dam removal and river restoration, Ecol. Eng., 28, 311–323, https://doi.org/10.1016/j.ecoleng.2006.08.003, 2006.
Goode, J. R., Luce, C. H., and Buffington, J. M.: Enhanced sediment delivery in a changing climate in semi-arid mountain basins: Implications for water resource management and aquatic habitat in the northern Rocky Mountains, Geomorphology, 139–140, 1–15, https://doi.org/10.1016/J.GEOMORPH.2011.06.021, 2012.
Goto, D., Hamel, M. J., Hammen, J. J., Rugg, M. L., Pegg, M. A., and Forbes, V. E.: Spatiotemporal variation in flow-dependent recruitment of long-lived riverine fish: Model development and evaluation, Ecol. Modell., 296, 79–92, https://doi.org/10.1016/j.ecolmodel.2014.10.026, 2015.
Hempel, S., Frieler, K., Warszawski, L., Schewe, J., and Piontek, F.: A trend-preserving bias correction – the ISI-MIP approach, Earth Syst. Dynam., 4, 219–236, https://doi.org/10.5194/esd-4-219-2013, 2013.
Hendry, K., Cragg-Hine, D., O'Grady, M., Sambrook, H., and Stephen, A.: Management of habitat for rehabilitation and enhancement of salmonid stocks, Fish. Res., 62, 171–192, https://doi.org/10.1016/S0165-7836(02)00161-3, 2003.
Hockley, F. A., Wilson, C. A. M. E., Brew, A., and Cable, J.: Fish responses to flow velocity and turbulence in relation to size, sex and parasite load, J. R. Soc. Interface, 11, 20130814, https://doi.org/10.1098/rsif.2013.0814, 2014.
IPCC: Climate Change 2013 – The Physical Science Basis, edited by: Stocker, T. F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S. K., Boschung, J., Nauels, A., Xia, Y., Bex, V., and Midgley, P. M., Cambridge University Press, Cambridge, UK and New York, NY, USA, 1535 pp., 2014.
Johnson, A. C., Acreman, M. C., Dunbar, M. J., Feist, S. W., Giacomello, A. M., Gozlan, R. E., Hinsley, S. A., Ibbotson, A. T., Jarvie, H. P., Jones, J. I., Longshaw, M., Maberly, S. C., Marsh, T. J., Neal, C., Newman, J. R., Nunn, M. A., Pickup, R. W., Reynard, N. S., Sullivan, C. A., Sumpter, J. P., and Williams, R. J.: The British river of the future: How climate change and human activity might affect two contrasting river ecosystems in England, Sci. Total Environ., 407, 4787–4798, https://doi.org/10.1016/J.SCITOTENV.2009.05.018, 2009.
Karthe, D., Kasimov, N., Chalov, S., Shinkareva, G., Malsy, M., Menzel, L., Theuring, P., Hartwig, M., Schweitzer, C., Hofmann, J., Priess, J., and Lychagin, M.: Integrating Multi-Scale Data For The Assessment Of Water Availability And Quality In The Kharaa–Orkhon–Selenga River System, Geogr. Environ. Sustain., 3, 65–86, https://doi.org/10.24057/2071-9388-2014-7-3-40-49, 2014.
Karthe, D., Chalov, S., Moreido, V., Pashkina, M., Romanchenko, A., Batbayar, G., Kalugin, A., Westphal, K., Malsy, M., and Flörke, M.: Assessment of runoff, water and sediment quality in the Selenga River basin aided by a web-based geoservice, Water Resour., 44, 399–416, https://doi.org/10.1134/S0097807817030113, 2017.
Lange, S.: EartH2Observe, WFDEI and ERA-Interim data Merged and Bias-corrected for ISIMIP (EWEMBI), GFZ Data Serv., https://doi.org/10.5880/pik.2016.004, 2016.
Moreido, V. M. and Kalugin, A. S.: Assessing possible changes in Selenga R. water regime in the XXI century based on a runoff formation model, Water Resour., 44, 390–398, https://doi.org/10.1134/S0097807817030149, 2017.
Motovilov, Y. G.: Hydrological simulation of river basins at different spatial scales: 1. Generalization and averaging algorithms, Water Resour., 43, 429–437, https://doi.org/10.1134/S0097807816030118, 2016.
Motovilov, Y. G., Gottschalk, L., Engeland, K., and Rodhe, A.: Validation of a distributed hydrological model against spatial observations, Agr. Forest Meteorol., 98–99, 257–277, https://doi.org/10.1016/S0168-1923(99)00102-1, 1999.
Nhiwatiwa, T., Dalu, T., and Brendonck, L.: Impact of irrigation based sugarcane cultivation on the Chiredzi and Runde Rivers quality, Zimbabwe, Sci. Total Environ., 587–588, 316–325, https://doi.org/10.1016/j.scitotenv.2017.02.155, 2017.
Papadaki, C., Soulis, K., Muñoz-Mas, R., Martinez-Capel, F., Zogaris, S., Ntoanidis, L., and Dimitriou, E.: Potential impacts of climate change on flow regime and fish habitat in mountain rivers of the south-western Balkans, Sci. Total Environ., 540, 418–428, https://doi.org/10.1016/J.SCITOTENV.2015.06.134, 2016.
Poff, N. L. and Zimmerman, J. K. H.: Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows, Freshw. Biol., 55, 194–205, https://doi.org/10.1111/j.1365-2427.2009.02272.x, 2010.
Poff, N. L., Allan, J. D., Bain, M. B., Karr, J. R., Prestegaard, K. L., Richter, B. D., Sparks, R. E., and Stromberg, J. C.: The Natural Flow Regime, Bioscience, 47, 769–784, https://doi.org/10.2307/1313099, 1997.
Richter, B. D., Baumgartner, J. V., Powell, J., and Braun, D. P.: A Method for Assessing Hydrologic Alteration within Ecosystems, Conserv. Biol., 10, 1163–1174, https://doi.org/10.1046/j.1523-1739.1996.10041163.x, 1996.
Roseman, E. F., Kennedy, G. W., Boase, J., Manny, B. A., Todd, T. N., and Stott, W.: Evidence of Lake Whitefish Spawning in the Detroit River: Implications for Habitat and Population Recovery, J. Great Lakes Res., 33, 397–406, https://doi.org/10.1016/S0380-1330(07)70064-3, 2007.
Schneider, C., Laizé, C. L. R., Acreman, M. C., and Flörke, M.: How will climate change modify river flow regimes in Europe?, Hydrol. Earth Syst. Sci., 17, 325–339, https://doi.org/10.5194/hess-17-325-2013, 2013.
Shrestha, R. R., Peters, D. L., and Schnorbus, M. A.: Evaluating the ability of a hydrologic model to replicate hydro-ecologically relevant indicators, Hydrol. Process., 28, 4294–4310, https://doi.org/10.1002/hyp.9997, 2014.
Sokolov, A. V. and Kalyagin, L. F.: General evaluation of the Baikal omul recruitment level, in State and problems of the artificial recruitment of the Baikal omul, IP Complex, Saint-Petersburg, Russia, 7–15, 2001.
Stagl, J. C. and Hattermann, F. F.: Impacts of Climate Change on Riverine Ecosystems: Alterations of Ecologically Relevant Flow Dynamics in the Danube River and Its Major Tributaries, Water (Switzerland), 8, 566, https://doi.org/10.3390/w8120566, 2016.
Statham, P. J.: Nutrients in estuaries – An overview and the potential impacts of climate change, Sci. Total Environ., 434, 213–227, https://doi.org/10.1016/J.SCITOTENV.2011.09.088, 2012.
Thompson, J. R., Laizé, C. L. R., Green, A. J., Acreman, M. C., and Kingston, D. G.: Climate change uncertainty in environmental flows for the Mekong River, Hydrol. Sci. J., 59, 935–954, https://doi.org/10.1080/02626667.2013.842074, 2014.
Törnqvist, R., Jarsjö, J., Pietroń, J., Bring, A., Rogberg, P., Asokan, S. M., and Destouni, G.: Evolution of the hydro-climate system in the Lake Baikal basin, J. Hydrol., 519, 1953–1962, https://doi.org/10.1016/j.jhydrol.2014.09.074, 2014.
Voronov, M. G.: Eco-biological basis for omul recruitment efficiency increase in the Selenga river in current conditions, Saint-Petersburg, Russia, 1993.
Wang, Y., Lei, X., Wen, X., Fang, G., Tan, Q., Tian, Y., Wang, C., and Wang, H.: Effects of damming and climatic change on the eco-hydrological system: A case study in the Yalong River, southwest China, Ecol. Indic., https://doi.org/10.1016/j.ecolind.2018.07.039, in press, 2019.
Wu, H., Chen, J., Xu, J., Zeng, G., Sang, L., Liu, Q., Yin, Z., Dai, J., Yin, D., Liang, J., and Ye, S.: Effects of dam construction on biodiversity: A review, J. Clean. Prod., 221, 480–489, https://doi.org/10.1016/j.jclepro.2019.03.001, 2019.